Генетические исследования на человеке имеют целый ряд вполне понятных ограничений. В связи с этим представляют интерес исследования генетических основ поведения у животных. Здесь можно применять методы селекции, получение инбредных линий, современные методы генной инженерии, избирательного выключения определенных генов, вызывать мутации и т.д. Инбредные линии, получаемые при длительном близкородственном скрещивании (не менее 20 поколений), представляют собой идентичных по генотипу животных, поэтому все отличия, которые можно наблюдать среди животных одной линии, связаны с воздействием среды.
Возникает вполне резонный вопрос: насколько данные, полученные на животных, допустимо переносить на человека?
Сходство геномов
В последнее время успешно идет расшифровка геномов человека и ряда животных. Если сравнить геном человека и, скажем, геном мыши, то становится очевидным наличие большого числа гомологичных генов. У обоих видов в геноме насчитывается приблизительно по 30 тыс. генов, причем для 99% генов мышей имеются соответствующие гомологичные гены у человека. Более того, большая часть генов мыши и человека занимают области хромосом с идентичными последовательностями генов (так называемые синтеничные области). 96% генов мыши найдено в синтеничных областях. Данные сравнительного генного анализа показывают, что в ходе эволюции млекопитающих линии, давшие, в конечном счете, человека и грызунов, разошлись сравнительно недавно — приблизительно 75 млн. лет тому назад. Основные отличия между этими видами обусловлены не столько числом генов и тем, какие это гены, сколько тем, как эти гены регулируются и взаимодействуют.
В любом случае не приходится сомневаться в том, что на молекулярном генном уровне между генами человека и мыши имеется значительная степень гомологии.
Сходство поведения
В то же время на уровне поведения картина намного сложнее, и вопрос о том, насколько уместно сравнение поведения этих двух видов, требует тщательного анализа в каждом конкретном случае.
Следует отметить, что у млекопитающих наблюдается значительное сходство в строении мозга и организации нейрональных систем, определяющих различные аспекты поведения. Следовательно, можно говорить о наличии общих нейроанатомических, нейрофизиологических механизмов, лежащих в основе поведения. Имеется большое сходство на уровне мотивационных процессов, таких как голод, жажда, страх, агрессия, половое поведение. Общими являются механизмы ряда форм обучения — привыкания, условно-рефлекторного обучения (классического и инструментального). Гомологичность (сходство) поведенческих механизмов проявляется и при реагировании на фармакологические вещества. Одни и те же фармакологические агенты, оказывающие влияние на нервную систему и психику человека, вызывают сходные реакции у млекопитающих. Это касается веществ, имеющих, например, седативное действие, или снимающих тревожные состояния и страх (транквилизаторов), психостимуляторов и т.п.
Все это заставляет полагать, что исследование генетики поведения животных может оказать огромную помощь в выяснении психогенетических основ поведения человека.
Примеры сравнительных генетических исследований человека и животных
В одной голландской семье было обнаружено характерное нарушение нормального поведения мужчин, которое заключалось во вспышках импульсивной агрессивности и импульсивном сексуальном поведении по отношению к женщинам. Наследование было слеплено с Х-хромосомой. Было выяснено, что наблюдавшийся синдром связан с точечной мутацией гена, кодирующего структуру фермента моноаминооксидазы А (МАОА).
Совершенно случайно в независимом исследовании подобный синдром был создан у мышей. Сходный фенотип у самцов был получен при создании линии мышей, предназначавшихся для иммунологических исследований (мышам попытались ввести ген интерферона). Эти сами^1 при групповом содержании демонстрировали повышенную агрессивность, что привело к появлению большого количества раненых животных. Они также проявляли частые попытки к захвату и спариванию с самками, которые не желали вступать в сексуальный контакт. При анализе оказалось, что при попытке ввести ген интерферона нарушили функцию гена моноаминооксидазы А (МАОА). Напомним, что моноаминооксидаза является важным ферментом, участвующим в регуляции функции катехоламинергических синапсов.
Этот пример хорошо иллюстрирует перспективность сравнительных психогенетических исследований.
Генетика поведения животных: насекомые
Приведем пример генетического анализа поведения, который довольно часто рассматривается в учебной литературе. Речь пойдет о пчелах и о заболевании под названием «американская личиночная гниль». Существует линия пчел, устойчивых к этой болезни, потому что в случае заболевания личинки пчелы немедленно распечатывают ячейку, в которой она находится, и удаляют ее из улья. Таким образом предупреждается распространение болезни, причем устойчивость к ней связана с характерным поведением! При скрещивании устойчивых к болезни пчел с неустойчивыми получают гибриды первого поколения (F1), которые не чистят ульи. Отсюда ясно, что аллель или аллели, обусловливающие этот тип поведения, рецессивные. Гибриды первого поколения F1 снова скрещивают с устойчивыми пчелами (так называемое анализирующее скрещивание — с рецессивными гомозиготными особями). В результате у потомства наблюдаются четыре варианта фенотипов в соотношении 1:1:1:1. Вот эти варианты:
- пчелы открывают ячейки, удаляют пораженные личинки;
- открывают ячейки, но не удаляют пораженные личинки;
- не открывают ячейки, но удаляют пораженные личинки, если ячейку откроет экспериментатор;
- не открывают ячейки, не удаляют пораженные личинки.
Таким образом, очевидно, что этот довольно сложный поведенческий акт контролируется генами всего в двух локусах. Один аллельный ген определяет действия по вскрытию ячейки, другой связан с удалением пораженной личинки.
В данном случае впечатляет тот факт, что довольно сложные действия могут контролироваться всего одним геном.
У плодовых мушек — дрозофил, которые долгие годы являлись излюбленным объектом генетиков — выявлено огромное количество мутаций, затрагивающих поведение. Так, мутация dunce приводит к нарушению способности к выработке условных рефлексов. Мутаций, так или иначе нарушающих обучение, известно несколько. Важно, что все эти дефекты связаны с нарушениями метаболизма так называемых вторичных мессенджеров (прежде всего циклической АМФ), играющих большую роль во внутриклеточной сигнализации и синаптической пластичности.
Есть мутации, приводящие к высокой или низкой половой активности, к избеганию определенных запахов, меняющие двигательную активность, вплоть до того, что есть мутация, определяющая, как дрозофила складывает крылья, — правое поверх левого или наоборот.
Иногда встречаются примеры весьма специфических отклонений в поведении. Так, при мутации fni (от fruitless — бесплодный) наблюдаются следующие нарушения полового поведения у самцов: они не ухаживают за самками, а ухаживают только за самцами, гомозиготными по данной мутации, и стимулируют нормальных самцов ухаживать за собой. Получилось нечто вроде модели формирования гомосексуального поведения.
Вообще складывается впечатление, что большинство поведенческих актов у дрозофил генетически предопределено во всех деталях.
Генетика поведения животных: млекопитающие
ДЛЯ поведения млекопитающих наряду с наличием безусловно-рефлекторного и инстинктивного поведения, опирающегося на врожденные механизмы, характерна относительно большая роль приобретенного индивидуального опыта. Этот факт уже подразумевает значительную роль средовых влияний в формировании особенностей поведения. Более того, при формировании фенотипа могут наблюдаться очень сложные взаимодействия генотипа и влияний внешней среды. Эту сложность хорошо иллюстрирует пример одной из мутаций у мышей. У нормальных мышей в соматосенсорной коре имеются характерные скопления нервных клеток, которые получили название «бочонки» (barrels). Каждый такой бочонок фактически является проекционной зоной для отдельной вибриссы. Вибриссы («усы»), расположенные на морде, у грызунов представляют собой очень важный сенсорный орган, играющий большую роль в поведении животного. Не случайно почти вся соматосенсорная кора у них занята такими «бочонками».
Была получена мутация brl (barrelless, т. е. «безбочоночные»), при которой нарушается формирование соматосенсорной коры и бочонки отсутствуют. Удалось идентифицировать на 11-й хромосоме соответствующий ген, и оказалось, что он кодирует структуру фермента аденилилциклазы типа I. Этот фермент синтезирует циклическую АМФ (цАМФ). Данное вещество является очень важным звеном во внутриклеточной сигнализации и играет большую роль в синаптической пластичности и процессах памяти. У мышей с этой мутацией действительно обнаруживается существенный дефицит памяти. Напомним, что у насекомых нарушения в метаболизме циклической АМФ также приводят к нарушению обучения.
То, что при данной мутации нарушается память, само по себе очень интересно, но какое отношение все это имеет к нарушению морфологической структуры коры? Оказалось, что самое непосредственное. При развитии соматосенсорной коры и при образовании в ней структур типа бочонков происходит процесс, который можно назвать сенсорным импринтингом. Сенсорный опыт, получаемый при стимуляции вибрисс, необходим для развития нормальной структуры коры и образования бочонков! Если этого опыта нет или нарушены механизмы его запоминания («импринтирование»), как в случае мутации brl, то наблюдаются существенные отклонения от нормальной морфологической структуры коры. Термин импринтинг в данном случае уместен, поскольку эти сенсорные воздействия играют решающую роль только в критический период развития, очень ограниченный во времени, а последствия этих воздействий сохраняются потом на всю жизнь.
Чем интересен данный пример? Он показывает, что генетическая программа, которая отвечает за развитие соматической сенсорной системы, обязательно учитывает при формировании ее механизмов сенсорные воздействия, которые поступают из окружающей среды.
В генетической программе запланирован учет реального сенсорного опыта! Иными словами, получается, что воздействие на нервную систему извне, т. е. то, что мы должны отнести к средовым воздействиям, является важнейшим условием правильной реализации генетической программы.
Похожие наблюдения были в свое время сделаны и в отношении других сенсорных систем, в частности зрительной. Так, например, в течение очень короткого времени (от нескольких часов до нескольких суток) после того, как у котят откроются глаза, на формирующиеся нейрональные механизмы зрения можно оказать серьезное влияние путем простого подбора сенсорных стимулов. Хьюбел и Визел показали, что характер сенсорной стимуляции в эти критические периоды может определять формирование механизмов нормального или патологического бинокулярного зрения или детекторные свойства нейронов зрительной коры, участвующих в распознавании формы объектов.
В вышеизложенных примерах средовые воздействия имеют настолько большое значение, что они фактически заложены во многие генетически запрограммированные формы поведения. Это касается не только формирования сенсорных систем. Рассмотрим, например, инстинкт гнездостроения у крыс. Считается, что это типично инстинктивное поведение, которое видоспецифично и наблюдается в отсутствие какого бы то ни было обучения. Если предоставить крысе перед рождением детенышей подходящий материал (обычно это тонкие полоски бумаги), то она успешно построит гнездо. Однако если содержать животное в условиях, при которых оно в течение всей жизни лишено возможности манипулировать мелкими предметами, то инстинкт гнездостроения уже не сможет реализоваться.
Хорошие примеры роли взаимодействия генетических и средовых факторов в формировании фенотипа можно почерпнуть из анализа противоречивости результатов, получаемых в разных лабораториях на генетически идентичных линиях животных. Довольно часто даже при выполнении одних и тех же тестов не удается воспроизвести результаты, полученные в другой лаборатории. Оказывается, что совсем небольшие отличия в методике эксперимента (скажем, постановка эксперимента утром или вечером) могут приводить к существенной разнице в результатах. Например, Максон сообщал, что влияние F-хромосомы на агонистическое поведение (нападения) в используемых им линиях исчезло, когда животных перевели в более чистую среду и стали подкислять воду для подавления роста бактерий (Мах-son, 1992). Иногда имеет значение даже то, кем проводится тестирование, — начинающим экспериментатором или опытным ученым.
Достаточно поучительный случай такого рода произошел с самим И. П. Павловым. Правда, здесь речь идет уже не о взаимодействии генотипа со средой. На конгрессе физиологов в 1923 г. было сделано сообщение, что при систематической выработке условных рефлексов из поколения в поколение у мышей наблюдается заметное снижение числа необходимых сочетаний для выработки прочного условного рефлекса. Если в первом поколении для выработки условного рефлекса потребовалось 300 сочетаний, то в четвертом поколении было достаточно только пяти сочетаний условного и безусловного стимула. Речь, таким образом, шла ни много ни мало о наследовании приобретенных признаков. Однако к чести исследователей они в конце концов разобрались, в чем дело. На самом деле речь шла о совершенствовании методики обучения, т. е. об обучении самого экспериментатора (а не мышей!). Исследование начинал неопытный исследователь, который постепенно набирал практические навыки работы с животными, из-за чего и наблюдался постоянный рост результатов от одного поколения к другому.
Исследования способности к обучению
Одним из самых важных свойств поведения животных является способность к обучению. Исследования на животных дают возможность провести селекционные эксперименты. Одним из первых на крысах такой эксперимент поставил Трайон. Он проводил селекцию по признаку обучаемости животных, которые должны были находить правильный путь к подкормке, помещенной в сложном 17-тупиковом лабиринте. Отбирались хорошо и плохо обучавшиеся животные, которые в дальнейшем скрещивались уже только между собой. Регулярная селекция дала очень быстрый результат — начиная с восьмого поколения показатели обучаемости у «умных» и «глупых» крыс (число ошибочных пробежек в лабиринте) уже не перекрывались. Селекция проводилась до 22-го поколения, в результате были получены две группы крыс — хорошо обучавшиеся (bright) и плохо — (dull). При одинаковых условиях выращивания и тестирования различия между этими группами обусловлены только отличиями в генотипе. В дальнейшем было получено множество линий, особенно у мышей, различавшихся по способности к различным формам обучения. Подобные линии были отобраны по способности обучаться в Т-образном лабиринте, по обучению активному и пассивному избеганию, плаванию в водном тесте Морриса. Иногда задача, которую выполняет животное, весьма сложна. Так, например, были получены линии мышей, хорошо и плохо обучавшиеся пищедобывательному двигательному условному рефлексу. Мыши получали подкрепление, когда они прыгали в ответ на световой или звуковой стимул на разные полочки (Корочкин, 1997). При этом можно отметить некоторые общие закономерности:
- обычно имеет место большое разнообразие признака в исходной популяции;
- хотя селекционный ответ может проявляться очень рано и разница между линиями обнаруживается уже через 2-3 поколения, для появления стабильных достоверных отличий между линиями требуется гораздо больше поколений (около 10-20).
Высокий разброс исходных значений признака и постепенное развитие селекционного ответа являются свидетельствами полигенной природы признака. Иными словами, проявление данного признака в фенотипе зависит от сравнительно большого числа генов. Подобным же образом обстоит дело с большинством черт поведения млекопитающих.
Существует еще одна проблема, связанная с опытами по селекции. Селекция проводится при тестировании какой-то одной определенной задачи. Естественно, возникает вопрос: насколько способность решать данную задачу коррелирует со способностью к другим видам обучения? Однозначного ответа на этот вопрос нет.
Например, когда стали более подробно изучать способность к обучению вообще на линиях крыс, полученных Трайоном (bright и dull), то выяснилось, что хорошо обучавшиеся (bright) быстрее обучаются пищедобывательному поведению, а крысы dull в свою очередь демонстрируют лучшие показатели в задачах на оборонительную реакцию. Таким образом, здесь проблема обучения может быть перенесена в плоскость мотивационных механизмов. Известно, что мотивация может исключительно сильно сказываться на результатах обучения.
Получается, что крысы линии bright сильнее мотивируются голодом, тогда как крысы dull сильнее мотивируются страхом в угрожающих ситуациях. Точно так же как мотивация, на успешность обучения могут влиять сенсорные способности, уровень двигательной активности, эмоциональность животных. Соответственно гены, влияющие на развитие данных качеств, могут оказывать воздействие и на обучение.
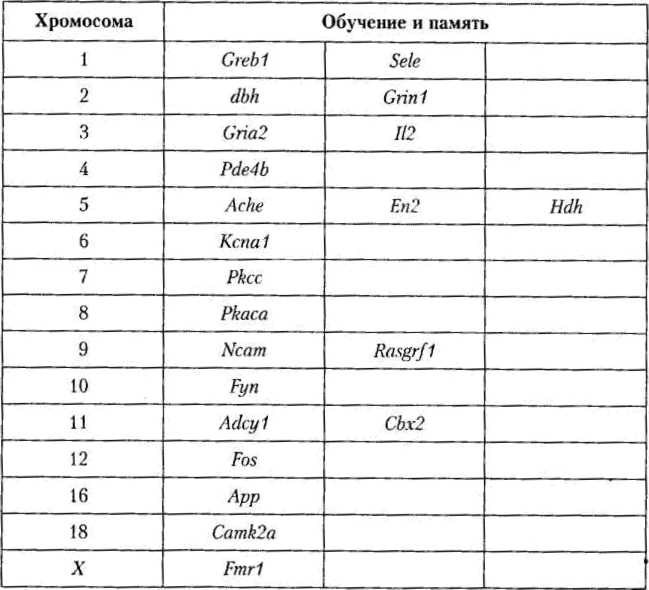
Однако некоторые линии демонстрируют различия и в более общих способностях к обучению. Так, мыши линии DBA/2J обучаются лучше, чем животные линии СВА, что подтверждается в целом ряде тестов: при пищевом подкреплении в лабиринте, в челночной камере при выработке условно-рефлекторной реакции активного избегания, при оперантном обучении. Значит, есть некие генетически определяемые свойства нервной системы, которые влияют на способность реализации различных типов обучения. Список мутаций, нарушающих обучение и память у мышей, быстро расширяется (табл. 4.1).
Таблица 4.1
Гены мыши, локализованные на определенных хромосомах и играющие важную роль в обучении и памяти (Wahlsten, 1999)
{kind=link}
У крыс Трайсна были отмечены и различия в характеристиках памяти что, безусловно, сказывалось на результатах тестирования. Так оказалось, что у крыс линии bright быстрее происходит консолидация — упрочение следов памяти, перевод их в устойчивую форму. Можно прибегнуть к воздействиям, нарушающим краткосрочную память, например, применить особую форму электрошока, вызывающую амнезию. Оказалось, что уже спустя 75 с после обучения элек-трошоковую амнезию не удается вызвать у крыс линии bright, тогда как на крыс линии dull электрошоковая процедура по-прежнему оказывает воздействие.
Разная скорость консолидации, по-видимому, определяет различия в успешности формирования навыков ориентации в лабиринте. Что произойдет, если крысам линии dull будет предоставлено время, достаточное для запоминания? Исследования показали, что когда интервалы между попытками составляли 30 с, крысы линии bright обучались намного быстрее крыс линии dull, как это и должно было быть. Но когда интервал увеличивали до 5 мин, разница в обучении между линиями существенно уменьшалась. Если же крысам вообще давали только 1 попытку в день, показатели обучаемости обеих линий становились одинаковыми. Скорость приобретения навыка и скорость консолидации может определяться разными механизмами!
Важный вывод: подбор условий обучения может уменьшить или даже свести на нет разницу в генетически обусловленных способностях.
В настоящее время получен целый ряд линий мышей, резко отличающихся по скорости консолидации памяти. Имеется линия (СЗН/ Не), у которой обучение возможно только при непрерывной тренировке без перерывов. Есть линия (DBA/2J), у которой обучение, наоборот, идет значительно успешнее при увеличении интервалов между отдельными сессиями тренировки. И наконец, выведена линия (BALB/c), для которой характер интервалов между экспериментальными сессиями не сказывается на результатах обучения. Данный подход создает, таким образом, уникальные возможности для изучения механизмов памяти.
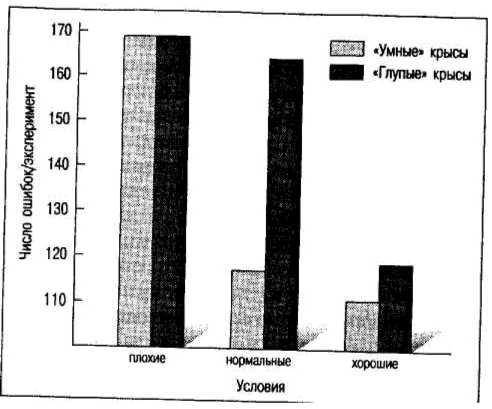
Еще одно направление исследований на животных — это выяснение влияний окружающей среды на формирование свойств поведения. Вновь вернемся к крысам линий bright и dull. Можно поставить эксперимент по выращиванию этих крыс в разных условиях. Одна группа (контрольная) выращивается в обычных условиях вивария. Для другой создается «обогащенная» среда — клетки больших размеров с раскрашенными стенками, наполненные различными предметами, зеркалами, мячами, трапами, лестницами, туннелями. Наконец, третьей группе предоставляется «обедненная» среда, где сильно ограничен приток сенсорных раздражителей и ограничены возможности поисковой и исследовательской активности. На рис. 4.1 обогащенная среда обозначена как «хорошие» условия, обедненная среда — как «плохие». Нормальные условия соответствуют контрольной группе.
Результаты обучения в лабиринте линий «умных» и «глупых» крыс, выросших в ухудшенных, обычных и улучшенных условиях (по Фогель, Мотульски, 1990)
{kind=link}
Результаты контрольной группы соответствуют ожиданию — крысы линии bright при обучении в лабиринте совершают намного меньше ошибок по сравнению с крысами линии dull. Однако для крыс, воспитанных в условиях обогащенной среды, эта разница практически сходит на нет, причем в основном за счет резкого уменьшения ошибок у «глупой» линии крыс. В случае воспитания в условиях обедненной среды разница между двумя линиями также исчезает, причем на этот раз в основном за счет резкого увеличения числа ошибок у «умной» линии крыс.
Здесь мы касаемся очень важной проблемы — существования мощных механизмов пластичности нервной системы, которые способны компенсировать весьма существенные дефекты. Многочисленные исследования по выращиванию крыс в обогащенной среде показали, что сравнительно быстро — за 25-30 дней — возникают весьма существенные морфологические отличия на уровне коры больших полушарий. У животных, содержавшихся в обогащенной среде, отмечается более толстая кора, большие размеры нейронов, на 10-20% увеличивается число дендритных отростков, приходящихся на один нейрон. Все это приводит к увеличению, на 20% числа синапсов, приходящихся на один нейрон. В конечном счете речь идет о миллиардах новых синапсов, что резко увеличивает возможности нервной системы. Особенно важен тот факт, что данный потенциал пластичности сохраняется практически все время.
Опыты на взрослых животных привели к аналогичным результатам.
Сходным образом обогащенная среда оказывает влияние и на развитие ребенка. Еще в 1940-е г. работы Спитца показали, что если ребенок воспитывается в обедненной среде, то это приводит к тяжелому отставанию в умственном развитии и нарушениям поведения, особенно социального. Спитц сравнил развитие новорожденных детей в приюте для найденышей и в яслях при нью-йоркской женской тюрьме. В обоих учреждениях поддерживались примерно одинаковые стандарты медицинского обслуживания, питания и гигиены. Отличия касались интенсивности воздействий окружающей среды как чисто сенсорного характера, так и социального. В приюте при тюрьме детские кровати были открытого типа, так что ребенок мог видеть других детей, приходящих к ним матерей, персонал, выполняющий различные действия. Кроме того, к детям ежедневно на ограниченный период допускались матери.
В отличие от этого заведения в доме для найденышей было гораздо меньше контактов с людьми (на одного человека персонала приходилось семь детей) и наблюдалось сильное ограничение сенсорных контактов из-за того, что конструкция кроватей не позволяла видеть других детей.
К чему привела такая сенсорная и социальная депривация новорожденных? Если в начале, в возрасте 4 месяцев, у детей из приюта для найденышей показатели по некоторым тестам были даже лучше, то спустя 8 месяцев (в конце первого года жизни) умственное и моторное развитие этих детей уже значительно отставало. У них быстро сформировался синдром, который Спитц назвал «госпитализмом»; в настоящее время его часто называют анаклитической депрессией. Эти дети практически не демонстрируют любопытства или веселости, всегда выглядят отстраненными, сильно подвержены всевозможным инфекциям (!).
В последующие два-три года последствия сенсорной и социальной депривации были еще более катастрофическими — к концу третьего года жизни из 26 детей только двое были способны говорить, и то лишь несколько слов.
Эксперименты по социальной депривации, проведенные на обезьянах, привели к очень похожим результатам. Выращивание новорожденных обезьян в изоляции от 6 месяцев до 1 года приводит к полной неспособности поддерживать нормальные социальные отношения в группе. Такие животные не взаимодействуют с другими обезьянами, не играют, не дерутся, отсутствуют проявления полового поведения. Обезьяны большую часть времени сидят скорчившись в углу клетки.
Здесь следует отметить очень важный момент — для возникновения синдрома изоляции также существует критический период в ходе развития организма. Точно такой же период изоляции после достижения 1,5-год овал ого возраста проходит у обезьян безо всяких последствий. Любопытно, что последующие исследования показали, что нормальное поведение можно восстановить, по крайней мере частично, если использовать некоторых обезьян, которые выполняют своеобразную роль «психотерапевтов». Такие обезьяны демонстрируют неослабевающую общительность, все время вовлекая животное с синдромом изоляции в социальное и агрессивное поведение до тех пор, пока изолянт не начинает отвечать.
Вышеприведенный пример еще раз указывает на наличие особых критических периодов на начальных этапах онтогенеза, когда воздействия среды имеют принципиально важное значение для формирования механизмов поведения. Даже сравнительно простые воздействия в этот период приводят к далеко идущим последствиям.
Рассмотрим еще один пример — так называемого хэндлинга. Это очень простая процедура, заключающаяся в том, что экспериментатор регулярно берет в руки детенышей, скажем крыс, на очень краткий период времени. Оказывается, что хэндлинг, особенно в первую неделю постнатального онтогенеза, приводит к целому ряду существенных изменений поведения. У животных, подвергавшихся хэнд-лингу, в дальнейшем наблюдается изменение реактивности по отношению к стрессорным факторам, повышается устойчивость к их воздействию, снижается эмоциональная реактивность, наблюдается ускорение полового созревания. Показано, что эти изменения, в частности связанные с устойчивостью к стрессорным воздействиям, сохраняются на всю жизнь и замедляют изменения, связанные со старением.
После хэндлинга наблюдается нормализация агрессивного поведения. В настоящее время имеются линии мышей с определенным уровнем агрессивности, варьирующим от очень высоких значений до низких. Под влиянием хэндлинга у линий мышей с исходно высокими значениями агрессивности она понижается, а у мышей со сниженной агрессивностью она, наоборот, повышается. Это еще один важный пример того, как поведение, «закодированное» в генотипе, можно регулировать подбором определенных внешних воздействий.
Проблема подбора осложняется наличием критических периодов чувствительности к воздействиям такого рода, часто очень непро-должительныим по времени. Кроме того, эффективность таких влияний достаточно сложно предсказать. С какой стати хэндлинг должен влиять на агрессивность? Эта связь изначально была далеко не так очевидна, особенно если учитывать довольно умеренный характер воздействия. Сейчас, когда уже многие подробности этих влияний выяснены, мы можем предложить ряд объяснений данного феномена, в частности изменение реакции на стресс в адренокортикальной системе, повышение чувствительности к определенным гормонам и т. п.
Исследования патологических форм поведения
На животных пытаются моделировать различные формы патологий человека, в том числе и патологического поведения. Это имеет большое значение как для изучения механизмов возникновения патологических состояний, так и для подбора средств лечения.
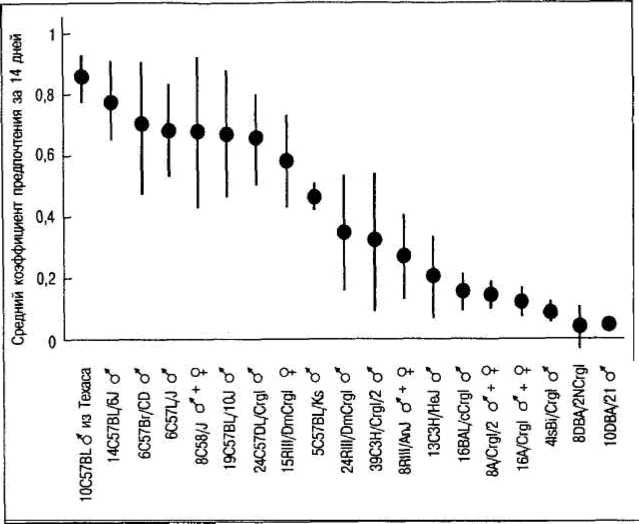
Так, при помощи селекции были получены линии мышей и крыс с различной степенью склонности к употреблению алкоголя (рис. 4.2). Особенно велико предпочтение алкоголя у мышей линии C57BL/6. Если таким животным предоставить свободный доступ к воде и раствору алкоголя (обычно не выше 20%), то они предпочитают потреблять алкоголь. Мыши линии DBA/2, напротив, практически не прикасаются к алкоголю. Эти исследования показали, что у линий крыс и мышей, предпочитающих алкоголь, наблюдается более высокая активность ферментов, принимающих участие в его метаболизме. Как известно, этиловый спирт в организме сначала превращается в уксусный альдегид под воздействием фермента алкогольде-гидрогеназы. На следующей стадии другой фермент — альдегиддегидрогеназа — переводит уксусный альдегид в уксусную кислоту. У крыс и мышей, предпочитающих алкоголь (например, у мышей линии C57BL), отмечается повышенная активность альдегиддегидрогеназы и низкий уровень уксусного альдегида в крови, равно как и пониженные значения содержания эндогенного этанола. Таким образом, получается, что высокая активность этого фермента связана с предпочтением алкоголя.
{kind=link}
Рис. 4.2. Различия в предпочтении алкоголя у ряда инбредных линий мышей. Коэффициент предпочтения равен отношению потребляемых алкоголя и воды. Представлены средние и стандартные отклонения (по Фогель, Мотульски, 1990)
Исследования показали, что и среди обычных линий, традиционно употребляемых в экспериментах, у части животных при предоставлении доступа к раствору этанола может развиться предпочтение алкоголя. При этом можно заранее определить будущих «алкоголиков» по длительности наркотического сна, вызываемого этанолом. Животные с высокой скоростью метаболизма алкоголя и соответственно непродолжительным сном оказываются способными к развитию стойкой алкогольной мотивации, тогда как долгоспящие редко начинают отдавать предпочтение алкоголю.
При развитии предпочтения алкоголя у животных можно оценить и роль средовых факторов. Оказалось, что всевозможные стрес-сорные ситуации, такие как конфликты, изоляция, неполноценное питание (например, диета с дефицитом витаминов), стимулируют потребление алкоголя, подобно тому, как это наблюдается и у человека.